Условно-рефлекторная деятельность коры больших полушарий. Физиология коры больших полушарий
Мозг это загадочный орган, который постоянно изучается учеными и остается до конца не исследованным. Система строения не простая и является сочетанием нейронных клеток, которые группируются в отдельные отделы. Кора головного мозга имеется у большинства животных и млекопитающих, но именно в человеческом организме она получила большего развития. Этому способствовала трудовая активность.
Почему мозг называют серым веществом или серой массой? Он сероватый, но в нем присутствует белый, красный и черные цвет. Серая субстанция представляет разные типы клеток, а белая нервную материю. Красный цвет это кровяные сосуды, а черный это меланин пигмент, который отвечает за окраску волос и кожи.
Строение мозга
Главный орган делится на пять основных частей. Первая часть продолговатая. Это продление спинного мозга, который контролирует связь с деятельностью тела и состоит из серой и белой субстанции. Вторая, средняя включает четыре бугорка, из которых два ответственные за слуховую, а два за зрительскую функцию. Третья, задняя включает мосток и церебеллум или мозжечок. Четвертая, буферная гипоталамус и таламус. Пятая, конечная, которая формирует два полушария.
Поверхность состоит из бороздочек и мозгов, покрытых оболочкой. Этот отдел составляет 80 % общего веса человека. Также мозг можно разделить на три части церебеллум, стволик и полушария. Он покрыт тремя слоями, которые предохраняют и питают основной орган. Это паутинный слой, в котором циркулирует мозговая жидкость, мягкий содержит кровяные сосуды, твердый близкий к мозгу и защищает его от повреждений.
Функции мозга

Мозговая деятельность включает основные функции серого вещества. Это чувствительные, зрительные, слуховые, обонятельные, осязательные реакции и моторные функции. Однако все главные центры управления находятся в продолговатой части, где координируется деятельность сердечно-сосудистой системы, защитных реакций и мышечной деятельности.
Двигательные пути продолговатого органа создают перекрещивание с переходом на противолежащую сторону. Это ведет к тому, что рецепторы сначала образуются в правой области, после чего поступают импульсы в левую область. Речь выполняется в больших полушариях мозга. Задний отдел отвечает за вестибулярный аппарат.
Как устроен наш мозг? Сколько нейронов в нем и каковы функции неокортекса? Современные ученые скрупулезно исследуют особенности нашего мозга и открывают все больше интересных подробностей.
Благодаря развитию высших нервных центров человек определяет себя и свое место в социуме, сознательно контролирует свое поведение и способен к адаптации в новой среде. Все эти преимущества связаны с функциями больших полушарий, которые мы рассмотрим.
Особенности мозга человека
Мозг человеческого вида весит приблизительно 1 кг 200 грамм - это средние показатели. Он состоит из 5 основных частей: это конечный, промежуточный, средний, задний и продолговатый мозг.

Большие борозды (углубления) разделяют 4 основные части лобную долю от теменной; а теменную - от затылочной; примыкает к трем другим. Последняя, пятая доля - островковая, которая находится в глубине латеральной ямки. Гармоничное взаимодействие всех нейронов обеспечивает рост и развитие нашей индивидуальности, наш характер и способности.
Можно выделить отдельную функцию больших полушарий - непрекращающееся развитие. Мозг человека все время развивается. Все, что индивидуум читает, видит, воспринимает, он буквально впитывает в себя. Особенно важна новая информация для детей до 2 лет, в это время их нейроны активно выстраивают связи на будущее.
Большие полушария. Строение и функции
В коре имеется от 14 до 17 млрд нейронов; а связей между клетками во много раз больше. Нейроны соединены синапсами. А помогают активировать связи различные нейромедиаторы - химические вещества, которые активируют рядом находящийся синапс.
Полушария мозга имеют особую структуру. Благодаря складкам, состоящим из борозд и извилин, площадь коры значительно увеличивается. По некоторым данным, общая площадь коры у среднестатистического человека - 2200 кв. см.
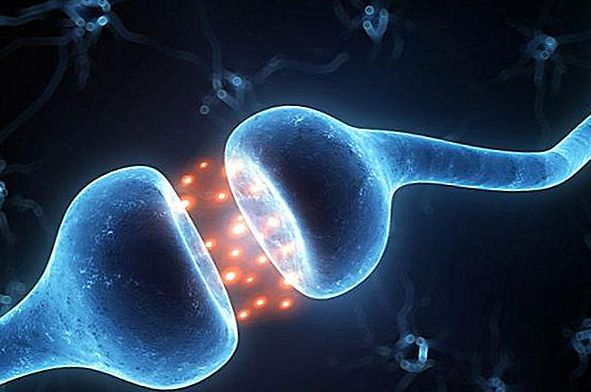
Под корой находится подкорка, или белое вещество мозга. Полушария между собой соединены мозолистым телом. А еще глубже находятся желудочки мозга - заполненные спинномозговой жидкостью пространства.
Кора состоит из слоев нервных нервных клеток, которые чередуются со слоями их ответвлений - аксонов. Всего насчитывается 6 слоев:
- молекулярный слой;
- наружный зернистый;
- наружный пирамидный - содержит преимущественно пирамидные нейроны;
- внутренний зернистый;
- внутренний пирамидный;
- слой веретеновидных нейронов.

Веретеновидные нейроны постепенно переходят в мозга. В коре происходят сознательные действия, формируется речь. В нижних глубинных частях под корой расположены центры бессознательных рефлексов и контроль внутренних органов и систем органов.
Зоны мозга
Чтобы понять функции больших полушарий головного мозга, нужно сначала разобрать их структуру. Полушария разделены условно на несколько центров, в которых проходят определенные психические и физиологические процессы. Эти центры не являются какими-то отдельными структурами. Все нейроны всех сетей постоянно взаимодействуют друг с другом. Это подтверждают многие исследователи.

Но все-таки можно выделить некоторые области в сером веществе мозга, которые более специализируются на отдельных задачах.
Зоны мозга нейрофизиологи выделяют следующие:
- Затылочная зона.
- Височная - отвечает за обоняние и вкус. Два эти чувства сильно взаимосвязаны.
- Зрительная зона. Тут расшифровываются сигналы, поступающие от глаз.
- Теменная - это так называемая зона кожно-мышечной чувствительности.
- Лобная доля - это сознательное поведение человека, его установки и трудовая деятельность. Задняя часть лобной доли - двигательный центр.
Функции больших полушарий мозга, как видим, распределены по зонам. Некоторые области имеют несколько функций. Например, руки связаны в больших полушариях с двумя зонами - двигательной и чувствительной.
И если при черепно-мозговой травме будет повреждена какая-либо из указанных областей, то функция этой зоны пострадает или совершенно пропадет. Восстановить утраченную функцию можно в том случае, если другая часть мозга - та, где находились нейроны, связанные с поврежденными тканями, сможет взять на себя всю работу утраченного центра.
Функции коры
Итак, каковы функции коры больших полушарий? Кора мозга отвечает за условные рефлексы, сформированные в процессе накопления опыта. Также в коре проходят все высшие психические процессы. Здесь сосредоточены зоны памяти, речи, мышления. Это более поздняя биологическая структура по сравнению с древним центральным мозгом, и она плохо изучена. Но известно, что наша личность и особенности характера, способность к усвоению и анализированию информации заложены именно в коре.
Большую роль играют в формировании навыков и привычек ассоциативные области. Можно сказать, утрируя информацию, что самая основная функция коры коры больших полушарий именно ассоциативная. Ведь на основе этих механизмов формируется и личность.
Ассоциативных областей 3:
- теменно-затылочно-височная;
- префронтальная ассоциативная;
- лимбическая.
Совместная работа этих центров обеспечивает всесторонний анализ поступающей извне информации. Без этих высших центров человек не смог бы целенаправленно выполнять работу.
Двигательная активность
Важнейшая функция больших полушарий - физическая активность. В передних отделах предцентральной извилины находится центр, где локализованы области проекции ступней и голеней. В средней части этой извилины находятся клетки, работающие с сигналами верхних конечностей, а самая глубокая часть предцентральной извилины отвечает за работу мышц лица.

Слаженная работа рецепторов проводящих нервных путей и этих мозговых центров обеспечивает нам ходьбу, работу руками и другую двигательную активность. Причем это все контролируется автоматически. Спортсмен ведь уже не думает, как согнуть ногу во время бега. Достаточно только дать сигнал старта сознательно.
Память и речь
В формировании памяти играют роль медиальная височная зона и гиппокамп. Однако они не являются тем местом, где накопленная информация хранится. Это скорее служебные зоны. Считается, что человек запоминает все, что видел или слышал когда-то. Основная проблема заключается в способности воспроизведения информации и ее перекодирования в слова.

Область речи - это граница височной и теменной зон. Причем у человека различают 2 зоны: отвечающий за речевое восприятие центр Вернике и за само произношение центр Брока.
Как лучше запомнить информацию?
Одна из функций больших полушарий, как мы теперь понимаем, - это запоминание и воспроизведение закодированной информации в словах. Если держать в мыслях и постоянно повторять одни и те же слова, то информация останется только в зоне речи и через несколько дней исчезнет.
Чтобы более глубоко запомнить информацию, необходимо применять образное мышление, ассоциируя каждое абстрактное понятие с яркими объектами.

В глубинной памяти у нас сохраняются только те аспекты реальности, которые связаны с яркими впечатлениями и сильными продолжительными эмоциями. А эмоции у нас "базируются" глубоко в белом веществе - в миндалевидном теле. Функции больших полушарий связаны с чисто сознательными намерениями запомнить.
Стрессы и депрессии ухудшают способность мозга запоминать что-либо. Начинать учить материал в беспокойном или раздражительном состоянии попросту бесполезно.
Вывод
Что можно сказать о функциях больших полушарий? Все центры мозга тесно взаимосвязаны. Говоря о конкретных областях, ученые подразумевают скопление нейронов, которые больше других взаимосвязанных сетей участвуют в том или ином психическом процессе.
Формирование памяти, способность говорить и думать словами - это самый сложный психический процесс. На это уходит большое количество энергии, и речью занято множество нервных клеток.
Кора больших полушарий связана непосредственно с сознательными процессами, а подкорка - с бессознательными, глубинными частями личности, которое Фрейд называл "Оно".
Раньше считалось, что высшие функции мозга человека осуществляются корой больших полушарий. Еще в прошлом веке было установлено, что при удаление коры у животных, они теряют способность к выполнению сложных актов поведения, обусловленных приобретенным жизненным опытом. Сейчас установлено, что кора не является высшим распределителем всех функций. Многие ее нейроны входят в состав сенсорных и двигательных систем среднего уровня. Субстратом высших психических функций являются распределительные системы ЦНС, в состав которых входит и подкорковые структуры, и нейроны коры. Роль любой области коры зависит от внутренней организации её спналтических связей, а также ее связей с другими образованиями ЦНС. Вместе с тем. у человека в процессе эволюции произошла кортиколизация всех, в том числе и жизненно важных висцеральных функций. Т.е. их подчинение коре. Она стала главной интегрирующей системой всей ЦНС. Поэтому в случае гибели значительной части нейронов коры у человека, его организм становится нежизнеспособным и погибает в результате нарушения гомеостаза (гипотермия мозга). Кори головного мозга состоит из шести слоев:
I. Молекулярный слой, самый верхний. Образован множеством восходящих дендритов пирамидных нейронов. Тел нейронов в нем мало. Этот слой пронизывают аксоны неспецифических ядер таламуса относящихся к ретикулярной формации. За счет такой структуры слой обеспечивает активацию всей коры.
2-Наружный зернистый слой. Формируется плотно расположенными мелкими нейронами, имеющими многочисленные синаптические контакты между собой. Благодаря этому наблюдается длительная циркуляция нервных импульсов. Это является одним из механизмов памяти.
3. Наружный пирамидный слой. Состоит из мелких пирамидных клеток. С помощью их и клеток второго слоя происходит образование межкортикальных связей, т.е. связей между различными областями коры.
4. Внутренний зернистый слой. Содержит звездчатые клетки, на которых образуют синапсы аксоны переключающих и ассоциативных нейронов таламуса. Сюда поступает вся информация от периферических рецепторов.
5. Внутренний пирамидный слой. Образован крупными пирамидными нейронами, аксоны которых образуют нисходящие пирамидные пути, направляющиеся в продолговатый и спинной мозг.
6. Слой полиморфных клеток. Аксоны его нейронов идут к таламусу.
Корковые нейроны образуют нейронные сети, включающие три основных компонента:
1. афферентные или входные волокна.
2.интернейроны
3. эфферентные - выходные нейроны. Эти компоненты образуют несколько уровней нейронных сетей.
1. микросети. Самый нижний уровень. Это отдельные межнейронные синапсы с их пре- и постсинаптическими структурами Синапс является сложным функциональным элементом, имеющим внутренние саморёгуляторные механизмы. Нейроны коры имеют сильно разветвленные дендриты. На них находится огромное количество шипиков в виде барабанных палочек. Эти шипики служат для образования входных синапсов. Корковые синапсы чрезвычайно"" чувствительны к внешним воздействиям. Например, лишение зрительных раздражений, путем содержания растущих животных в темноте, приводит к значительному уменьшению синапсов в зрительной коре. При болезни Дауна синапсов в коре также меньше, чем в норме. Каждый шипик образующий синапс, выполняет роль преобразователя сигналов идущих к нейрону.
2. Локальные сети. Новая кора слоистая структура, слои которой образованы локальными нейронными сетями. К ней через таламус и обонятельный мозг, могут приходить импульсы от всех периферических рецепторов. Входные волокна проходят через все слои, образуя синапсы с их нейронами. В свою очередь, коллатерали входных волокон и интернейроны этих слоев образуют локальные сети на каждом уровне коры. Такая структура коры обеспечивает возможность обработки, хранения и взаимодействия различной информации. Кроме того, в коре имеется несколько типов выходных нейронов. Практически каждый ее слой дает выходные волокна, направляющиеся к другим слоям или отдаленным участкам коры.
3. Корковые колонки. Входные и выходные элементы с интернейронами образуют вертикальные корковые колонки пли локальные модули. Они проходят через все слои коры. Их диаметр составляет 300-500 мкм. Образующие эти колонки нейроны концентрируются вокруг таламо-кортикального волокна, несущего определенный вид сигналов. В колонках имеются многочисленные межнейронные связи. Нейроны 1-5 слоев колонок обеспечивают восприятие и переработку поступающей информации. Нейроны 5-6 слоя образуют эфферентные пути коры. Соседние колонки также связаны между собой. При этом возбуждение одной сопровождается торможением соседних. В определенных областях коры сосредоточены колонки, выполняющие однотипную функцию. Эти участки называются цитоархитектоническими полями. В коре человека их 53. Поля делят на первичные, вторичные, третичные.
Первичные обеспечивают обработку определенной сенсорной информации.
Вторичные и третичные взаимодействие сигналов разных сенсорных систем. В частности, первичное соматосенсорное поле, к которому идут импульсы от всех кожных рецепторов (тактильных, температурных, болевых) находится в области центральной задней извилины. Больше всего места в коре занимает представительство губ, лица, кистей рук. Поэтому при поражениях этой зоны изменяется чувствительность соответствующих участков кожи. Представительство проприорецепторов мышц и сухожилий, т.е. моторная кора занимает переднюю центральную извилину. Импульсы от проприорецепторов нижних конечностей идут к верхней части извилины. От мышц туловища к средней части. От мускулатуры головы и шеи к ее нижней части. Наибольшую площадь этого поля также занимает представительство мускулатуры губ, языка, кистей и лица.
Импульсы от рецепторов глаза поступают в затылочные области коры около шпорной борозды. Поражение первичных полей приводит к корковой слепоте, а вторичных и третичных - потере зрительной памяти. Слуховая область коры расположена в верхней височной извилине и поперечной извилине Гешля. При поражении первичных полей зоны развивается корковая глухота. Периферических - трудности в различении звуков. В задней трети верхней височной извилины левого полушария находится сенсорный центр речи - центр Вернике. При его патологических изменениях теряется способность к пониманию речи. Двигательный центр речи - центр Брока, располагается в нижней лобной извилине левого полушария. Нарушения в этой части коры приводят к потере способности произносить слова.
Функциональная асимметрия полушарий.
Передний мозг образован двумя полушариями, которые состоят из одинаковых долей. Однако они играют разную функциональную роль. Впервые различия между полушариями описал 1863 г. невропатолог Поль Брэка. обнаруживший, что при опухолях левой лобной доли теряется способность к произношению речи. В 50-х годах XX века Р.Сперри и М.Газзанига исследовали больных, у которых с целью прекращения эпилептических припадков была произведена перерезка мозолистого тела. В нем проходят комиссуральные волокна, связывающие полушария. Умственные способности у людей с расщепленным" мозгом не изменяются. Но с помощью специальных тестов обнаружено, что функции полушарий отличаются. Например, если предмет находится в поле зрения правого глаза, то зрительная информация поступает в левое полушарие, то такой больной может назвать его, описать его свойства. прочитать или написать текст.
Если же предмет попадает в поле зрения левого глаза, то пациент даже не может назвать его и рассказать о нем. Он не может читать этим глазом. Таким образом, левое полушарие является доминирующим в отношении сознания, речи, счета, письма, абстрактного мышления, сложных произвольных движений. С другой, стороны, хотя правое полушарие не имеет выраженных речевых функций, оно в определенной степени способно понимать речь и мыслить абстрактно. Но в значительно большей мере, чем левое, оно обладает механизмами сенсорного распознавания предметов образной памяти. Восприятие музыки целиком является функцией правого полушария. Т.е. правое полушарие отвечает за неречевые функции, т.е. анализ сложных зрительных и слуховых образов, восприятие пространства, формы. Каждое полушарие изолированно принимает, перерабатывает и хранит информацию. Они обладают собственными ощущениями, мыслями, эмоциональными оценками событий. Левое полушарие обрабатывает информацию аналитически, т.е. последовательно, а правое одномоментно, интуитивно. т.е. полушария используют разные способы познания. Вся система образования в мире направлена на развитие левого полушария, т.е. абстрактного мышления, а не интуитивного. Несмотря на функциональную асимметрию, в норме полушария работают совместно, обеспечивая все процессы человеческой психики.
Пластичность коры.
Некоторые ткани сохраняют способность к образованию новых клеток из клеток-предшественников в течение всей жизни. Это клетки печени, кожи энтероциты. Нервные клетки не обладают такой способностью. Однако у них сохраняется способность к образованию новых отростков и синапсов т.е каждый нейрон способен при повреждении отростка образовывать новые. Восстановление отростков может происходить двумя путями: путем формирования нового конуса роста и образования коллатералей. Обычно росту нового аксона препятствует возникновение, глиального рубца. Но несмотря на это новые синаптические контакты образуются коллатералям и поврежденного аксона. Наиболее высока пластичность нейронов коры. Любой ее нейрон запрограммирован на то, что при его повреждении он активно пытается восстановить утраченные связи. Каждый нейрон вовлечен, а конкурентную борьбу с другими за образование синаптических контактов. Это служит основой пластичности нейронных корковых сетей. Установлено, что при удалении мозжечка нервные пути, идущие к нему, начинают прорастать в кору. Если в интактный мозг пересадить участок мозга другого животного, то нейроны этого кусочка ткани образуют многочисленные контакты с нейронами мозга реципиента.
Пластичность коры проявляется как в нормальных условиях. Например, при образовании новых межкортикальных связей в процессе обучения, так и при патологии. В частности, утраченные при поражении участка коры функции берут на себя ее соседние поля или другое полушарие. Даже при поражении обширных областей коры вследствие кровоизлияния, их функции начинают выполнять соответствующие области противоположного полушария.
Элгктроэнцефалография. Ее значение для экспеперементальных исследований и клиники.
электроэнцефалография (ЭЭГ - это регистрация электрической активности мозга с поверхности кожи головы. Впервые ЭЭГ человека зарегистрировал в 1929 г. немецкий психиатр Г.Бергер. При снятии ЭЭГ на кожу накладывают электроды, сигналы от которых усиливаются и подаются на осциллограф и пишущее устройство. В норме регистрируются следующие типы спонтанных колебаний:
1. а-ритм. Это волны с частотой 8-13 Гц. Наблюдается в состоянии бодрствования, полного покоя и при закрытых глазах. Если человек открывает глаза а-ритм сменяется р-ритмом. Это явление называется блокадой а-ритма.
2. В-ритм, Его частота от 14 до 30 Гц. Наблюдается при деятельном состоянии мозга и читается по мере повышения интенсивности умственной работы.
3. (гама) - ритм. Колебания с частотой 4-8 Гц. Регистрируется во время засыпания.
поверхностного сна и неглубоком наркозе.
4. (сигма) - ритм. Частота 0,5-3,5 Гц. Наблюдается при глубоком сне и наркозе.
Чем ниже частота ритмов ЭЭГ, тем больше их амплитуда. Помимо эти основных ритмов регистрируются и другие ЭЭГ феномены. Например, по мере углубления сна появляются сонные веретена. Это периодическое увеличение частоты и амплитуды тета- ритма. При ожидании команды к действию возникает отрицательная Е-волна ожидания и т.д.
В эксперименте ЭЭГ используют для определения уровня активности мозга, а в клинике для диагностики эпилепсии (особенно скрытых форм), а также для выявление смерти мозга (кора живет 3-5 мин, стволовые нейроны 7-10, сердце 90. почки 150).
Тема: Физиология ЦНС
Лекция №6– Общая характеристика головного мозга. Физиология продолговатого, среднего, промежуточного мозга, мозжечка, лимбической системы и коры больших полушарий.
Цель – Дать представление о роли различных отделов головного мозга в интегративной деятельности человека.
Головной мозг состоит из продолговатого мозга (его вместе с мостом называют задним мозгом), среднего и промежуточного мозга, мозжечка, базальных ядер, лимбической системы и коры больших полушарий. Каждый из них выполняет свою важную функцию, но в целом обеспечивает физиологические функции внутренних органов, скелетной мускулатуры и осуществление деятельности организма как единого целого.
Продолговатый мозг и варолиев мост – их относят к заднему мозгу, который является частью ствола мозга. Задний мозг осуществляет сложную рефлекторную деятельность и служит для соединения спинного мозга с вышележащими отделами головного мозга. В срединной его области расположены задние отделы ретикулярной формации, оказывающие неспецифические тормозные влияния на спинной и головной мозг.
Через продолговатый мозг проходят восходящие пути от рецепторов слуховой и вестибулярной чувствительности. Функции нейронов вестибулярных ядер продолговатого мозга разнообразны. Одна часть их реагирует на перемещение тела (например, при горизонтальных ускорениях в одну сторону они увеличивают частоту разрядов, а при ускорениях в другую сторону уменьшают их). Другая часть предназначена для связи с моторными системами. Эти вестибулярные нейроны, повышая возбудимость мотонейронов спинного мозга и нейронов двигательной зоны коры больших полушарий, позволяют регулировать двигательные акты в соответствии с вестибулярными влияниями.
В продолговатом мозгу оканчиваются афферентные нервы, несущие информацию от рецепторов кожи и мышечных рецепторов. Здесь они переключаются на другие нейроны, образуя путь в таламус и далее в кору больших полушарий. Восходящие пути кожно-мышечной чувствительности (как и большая часть нисходящих кортико-спинальных волокон) перекрещиваются на уровне продолговатого мозга.
В продолговатом мозгу и варолиевом мосту находится большая группа черепно-мозговых ядер (от V до XII пары), иннервирующих кожу, слизистые оболочки, мускулатуру головы и ряд внутренних органов (сердце, легкие, печень). Совершенство этих рефлексов обусловлено наличием большого количества нейронов, образующих ядра и соответственно большого числа нервных волокон. Так, только в одном нисходящем корешке тройничного нерва, проводящем болевую, температурную и тактильную чувствительность от головы, содержится во много раз больше волокон, чем в спинно-таламическом пути, содержащем волокна, идущие от болевых и температурных рецепторов остальной части тела.
На дне IV желудочка в продолговатом мозгу находится жизненно важный дыхательный центр, состоящий из центров вдоха и выдоха и пнеймотаксического отдела. Его составляют мелкие нервные клетки, посылающие импульсы к дыхательным мышцам через мотонейроны спинного мозга. В непосредственной близости расположены сердечный и сосудо -двигательный центры. Они регулируют деятельность сердца и состояние сосудов. Функции этих центров взаимосвязаны. Ритмические разряды дыхательного центра изменяют частоту сердечных сокращений, вызывая дыхательную аритмию - учащение сердцебиений на вдохе и замедление их на выдохе.
В продолговатом мозгу находится ряд рефлекторных центров, связанных с процессами пищеварения. Это группа центров моторных рефлексов (жевания, глотания, движений желудка и части кишечника), а также секреторных (слюноотделение, выделение пищеварительных соков желудка, поджелудочной железы и др.). Кроме того, здесь находятся центры некоторых защитных рефлексов: чихания, кашля, мигания, слезоотделения, рвоты.
Продолговатый мозг играет важную роль в осуществлении двигательных актов и в регуляции тонуса скелетных мышц. Влияния, исходящие из вестибулярных ядер продолговатого мозга, усиливают тонус мышц-разгибателей, что важно для организации позы.
Неспецифические отделы продолговатого мозга, наоборот, оказывают угнетающее влияние на тонус скелетных мышц, снижая его и в мышцах-разгибателях. Продолговатый мозг участвует в осуществлении рефлексов поддержания и восстановления позы тела, так называемых установочных рефлексов.
Средний мозг. Через средний мозг, являющийся продолжением ствола мозга, проходят восходящие пути от спинного и продолговатого мозга к таламусу, коре больших полушарий и мозжечку.
В состав среднего мозга входят четверохолмия, черная субстанция и красное ядро . Срединную его часть занимает ретикулярная формация, нейроны которой оказывают мощное активирующее влияние на всю кору больших полушарий, а также на спинной мозг.
Передние бугры четверохолмия представляют собой первичные зрительные центры, а задние бугры-первичные слуховые центры. Ими осуществляются также реакции, являющимися компонентами ориентировочного рефлекса при появлении неожиданных раздражителей. В ответ на внезапное раздражение происходит поворот головы и глаз в сторону раздражителя, а у животных-настораживание ушей. Этот рефлекс (по И. П. Павлову, рефлекс «Что такое?») необходим для подготовки организма к своевременной реакции на любое новое воздействие. Он сопровождается усилением тонуса мыщц-сгибателей (подготовка к двигательной реакции) и изменениями вегетативных функций (дыхание, сердцебиения).
Средний мозг играет важную роль в регуляции движений глаз. Управление глазодвигательным аппаратом осуществляют расположенные в среднем мозгу ядра блокового (IV) нерва, иннервирующего верхнюю косую мышцу глаза, и глазодвигательного (III) нерва, иннервирующего верхнюю, нижнюю и внутреннюю прямые мышцы нижнюю косую мышцу и мышцу, поднимающую веко, а также расположенное в заднем мозгу ядро отводящего (VI) нерва, иннервирующего наружную прямую мышцу глаза. С участием этих ядер осуществляются поворот глаза в любом направлении, аккомодация глаза, фиксация взгляда на близких предметах путем сведения зрительных осей, зрачковый рефлекс (расширение зрачков в темноте и сужение их на свету).
У человека при ориентации во внешней среде ведущим является зрительный анализатор, поэтому особое развитие получили передние бугры четверохолмия (зрительные подкорковые центры). У животных с преобладанием слуховой ориентации (собака, летучая мышь), наоборот, в большей степени развиты задние бугры (слуховые подкорковые центры).
Черная субстанция среднего мозга имеет отношение к рефлексам жевания и глотания, участвует в регуляции тонуса мышц (особенно при выполнении мелких движений пальцами рук).
В среднем мозгу важные функции осуществляет красное ядро. О возрастании роли этого ядра в процессе эволюции свидетельствует резкое увеличение его размеров по отношению к остальному объему среднего мозга. Красное ядро тесно связано с корой больших полушарий, ретикулярной формацией ствола, мозжечком и спинным мозгом.
От красного ядра начинается руброспинальный путь к мотонейронам спинного мозга. С его помощью осуществляется регуляция тонуса скелетных мышц, происходит усиление тонуса мышц-сгибателей. Это имеет большое значение как при поддержании позы в состоянии покоя, так и при осуществлении движений. Импульсы, приходящие в средний мозг от рецепторов сетчатки глаза и от проприорецепторов глазодвигательного аппарата, участвуют в осуществлении глазодвигательных реакций, необходимых для ориентации в пространстве, выполнении точных движений. В опыте при перерезке мозга ниже красного ядра происходит возбуждение мышц – расгибателей и торможение мышц – сгибателей, что характеризуется определенной позой, называемой децеребрационной ригидностью.
Промежуточный мозг. В состав промежуточного мозга, который является передним концом ствола мозга, входят зрительные бугры - таламус и подбугровая область - гипоталамус.
Таламус представляет собой важнейшую «станцию» на пути афферентных импульсов в кору больших полушарий.
Ядра таламуса подразделяют на специфические и неспецифические.
К специфическим относят переключательные (релейные) ядра и ассоциативные. Через переключательные ядра таламуса передаются афферентные влияния от всех рецепторов тела. Это так называемые специфические восходящие пути. Они характеризуются соматотопической организацией. Особенно большое представительство в таламусе имеют эфферентные влияния, поступающие от рецепторов лица и пальцев рук. От таламических нейронов начинается путь к соответствующим воспринимающим областям коры - слуховым, зрительных и др. Ассоциативные ядра непосредственно не связаны с периферией. Они получают импульсы от переключающих ядер и обеспечивают их взаимодействие на уровне таламуса, т. е. осуществляют подкорковую интеграцию специфических влияний. Импульсы от ассоциативных ядер таламуса поступают в ассоциативные области коры больших полушарий, где участвуют в процессах высшего афферентного синтеза.
Помимо этих ядер, в таламусе имеются неспецифические ядра, которые могут оказывать как активирующее, так и тормозящее влияние на кору.
Благодаря обширным связям таламус играет важнейшую роль в жизнедеятельности организма. Импульсы, идущие от таламуса в кору, изменяют состояние корковых нейронов и регулируют ритм корковой активности. Между корой и таламусом существуют кольцевые кортико-таламические взаимосвязи, лежащие в основе образования условных рефлексов. С непосредственным участием таламуса происходит формирование эмоций человека. Таламусу принадлежит большая роль в возникновении ощущений, в частности ощущения боли.
Подбугровая область (гипоталамус) расположена под зрительными буграми и имеет тесные нервные и сосудистые связи с прилежащей железой внутренней секреции-гипофизом. Здесь расположены важные вегетативные нервные центры, регулирующие обмен веществ в организме, обеспечивающие поддержание постоянства температуры тела (у теплокровных) и другие вегетативные функции.
Участвуя в выработке условных рефлексов и регулируя вегетативные реакции организма, промежуточный мозг играет очень важную роль в двигательной деятельности, особенно при формировании новых двигательных актов и выработке двигательных навыков.
Базальные ядра – так называют группу ядер серого вещества, расположенных непосредственно под полушариями большого мозга. К ним относятся парные образования: хвостатое тело и скорлупа, составляющие вместе полосатое тело (стриатум), и бледное ядро (паллидум). Базальные ядра получают сигналы от рецепторов тела через зрительные бугры. Эфферентные импульсы подкорковых ядер направляются к нижележащим центрам экстрапирамидной системы. Подкорковые узлы функционируют в единстве с корой больших полушарий, промежуточным мозгом и другими отделами мозга. Это обусловлено наличием кольцевых связей между ними. Через эти подкорковые ядра могут соединять между собою разные отделы коры больших полушарий, что имеет большое значение при образовании условных рефлексов. Совместно с промежуточным мозгом подкорковые ядра участвуют в осуществлении сложных безусловных рефлексов: оборонительных, пищевых и др.
Представляя собой высший отдел мозгового ствола, базальные ядра объединяют деятельность нижележащих образований, регулируя мышечный тонус и обеспечивая необходимое положение тела во время физической работы. Бледное ядро выполняет моторную функцию. Оно обеспечивает проявление древних автоматизмов - ритмических рефлексов. С его деятельностью связано также выполнение содружественных (например, движения туловища и рук при ходьбе), мимических и других движений.
Полосатое тело оказывает на двигательную деятельность тормозящее, регулирующее влияние, угнетая функции бледного ядра, а также моторкой области коры больших полушарий. При заболевании полосатого тела возникают непроизвольные беспорядочные сокращения мышц (гиперкинезы). Они обусловливают некоординированные толчкообразные движения головы, рук и ног. Нарушения возникают также в чувствительной сфере - понижается болевая чувствительность, расстраиваются внимание и восприятие.
В настоящее время выявлено значение хвостатого тела в самооценке поведения человека. При неправильных движениях или умственных операциях из хвостатого ядра в кору больших полушарий поступают импульсы, сигнализирующие об ошибке.
Мозжечок. Это - надсегментарное образование, не имеющее непосредственной связи с исполнительными аппаратами. Мозжечок входит в состав экстрапирамидной системы. Он состоит из двух полушарий и червя, находящегося между ними. Наружные поверхности полушарий покрыты серым веществом - корой мозжечка, а скопления серого вещества в белом веществе образуют ядра мозжечка.
Мозжечок получает импульсы от рецепторов кожи, мышц и сухожилий через спинно-мозжечковые пути и через ядра продолговатого мозга (от спинно-бульбарного пути). Из продолговатого мозга в мозжечок поступают также вестибулярные влияния, а из среднего мозга-зрительные и слуховые. Корково-мосто-мозжечковый путь связывает мозжечок с корой больших полушарий. В коре мозжечка представительство различных периферических рецепторов имеет соматотопическую организацию. Кроме того, наблюдается упорядоченность связей этих зон с соответствующими воспринимающими областями коры. Так, зрительная зона мозжечка связана со зрительной зоной коры, представительство каждой группы мышц в мозжечке - с представительством одноименных мышц в коре и т. д. Такое соответствие облегчает совместную деятельность мозжечка и коры в управлении различными функциями организма.
Эфферентные импульсы от мозжечка поступают к красным ядрам ретикулярной формации, продолговатому мозгу, таламусу, коре и подкорковым ядрам.
Мозжечок участвует в регуляции двигательной деятельности. Электрические раздражения поверхности мозжечка вызывают движения глаз, головы и конечностей, которые отличаются от корковых моторных эффектов тоническим характером и большой длительностью. Мозжечок регулирует изменение и перераспределение тонуса скелетных мышц, что необходимо для организации нормальной позы и двигательных актов.
Функции мозжечка изучались в клинике при его поражениях у человека, а также у животных путем удаления (экстирпации мозжечка) (Л. Лючиани, Л. А. Орбели). В результате выпадения функций мозжечка возникают двигательные расстройства: атония- резкое падение и неправильное распределение тонуса мышц, астазия - невозможность сохранения неподвижного положения, непрерывные качательные движения, дрожание головы, туловища и конечностей, астения - повышенная утомляемость мышц, атаксия - нарушение координированных движений, походки и др.
Мозжечок оказывает влияние также на ряд вегетативных функций, например желудочно-кишечного тракта, на уровень кровяного давления, на состав крови.
Таким образом, в мозжечке происходит интеграция самых различных сенсорных влияний, в первую очередь проприоцептивных и вестибулярных. Мозжечок даже ранее считали центром равновесия и регуляции мышечного тонуса. Однако его функции, как оказалось, гораздо обширнее-охватывают также регуляцию деятельности вегетативных органов. Деятельность мозжечка протекает в непосредственной связи с корой больших полушарий, под ее контролем.
Функции ретикулярной формации. Различают два основных типа влияния неспецифической системы на работу других нервных центров - активирующее и тормозящее влияния. Оба они могут адресоваться как вышележащим центрам (восходящие влияния), так и нижележащим (нисходящие влияния).
Восходящие влияния. В опытах на животных было показано, что из сетевидного образования среднего мозга исходит мощное активирующее влияние на кору больших полушарий. Электрические раздражения этих отделов неспецифической системы через вживленные электроды вызывали пробуждение спящего животного. У бодрствующего животного подобное раздражение повышало уровень корковой активности, усиливало внимание к внешним сигналам и улучшало их восприятие.
Нисходящие влияния. Все отделы неспецифической системы оказывают помимо восходящих значительные нисходящие влияния. Отделы ствола мозга регулируют (активируют или угнетают) активность нейронов спинного мозга и проприорецепторов мышц (мышечных веретен). Эти влияния совместно с воздействиями из экстрапирамидной системы и мозжечка играют большую роль в регуляции тонуса мышц и обеспечении позы человека. Непосредственные команды к осуществлению движений и влияния, формирующие перестройки тонуса мышц, передаются по специфическим путям. Однако неспецифические влияния позволяют существенно изменить протекание этих реакций. При усилении активирующих воздействий из ретикулярной формации среднего мозга на нейроны спинного мозга увеличивается амплитуда производимых движений и повышается тонус скелетных мышц. Включение этих влияний при некоторых эмоциональных состояниях помогает повысить эффективность двигательной деятельности человека и выполнить значительно большую работу, чем в обычных условиях.
Возникновение эмоций, а также поведенческие реакции связывают с деятельностью лимбической системы, в которую входят некоторые подкорковые образования и участки коры. Корковые отделы лимбической системы, представляющие ее высший отдел находятся на нижних и внутренних поверхностях больших полушарий (поясная извилина, гиппокамп и др.). К подкорковым структурам лимбической системы относят также грушевидную долю, обонятельную луковицу и тракт, миндалевидное ядро, гипоталамус, некоторые ядра таламуса, среднего мозга и ретикулярной формации. Между всеми этими образованиями имеются тесные прямые и обратные связи образующие «лимбическое кольцо».
Лимбическая система участвует в самых разнообразных проявлениях деятельности организма. Она формирует положительные и отрицательные эмоции со всеми двигательными, вегетативными и эндокринными их компонентами (изменением дыхания, сердцебиения кровяного давления, деятельности желез внутренней секреции, скелетных и мимических мышц и др.). От нее зависит эмоциональная окраска психических процессов и изменения двигательной активности. Она создает мотивацию поведения (определенную предрасположенность ). Возникновение эмоций имеет «оценочное влияние» на деятельность специфических систем, так как, подкрепляя определенные способы действий, пути решения поставленных задач, они обеспечивают избирательный характер поведения в ситуациях со многими выборами. Области коры, относящиеся к лимбической системе (нижние и внутренние части коры), обеспечивают эмоциональную окраску движений и управляют вегетативными реакциями организма при работе.
Лимбическая система участвует в формировании ориентировочных и условных рефлексов. Благодаря центрам лимбической системы могут вырабатываться даже без участия других отделов коры оборонительные и пищевые условные рефлексы. При поражениях этой системы затрудняется упрочение условных рефлексов, нарушаются процессы памяти, теряется избирательность реакций и отмечается неумеренное их усиление (чрезмерно повышенная двигательная активность и т. д.). Известно, что так называемые психотропные вещества, изменяющие нормальную психическую деятельность человека, действуют именно на структуры лимбической системы. Таким образом, лимбическая система задает общий контекст поведения, в зависимости от условий, переводя в нужное предрасположенное состояние- эмоцию. Направленность эмоции (положительная или отрицательна) и определяет вид формирующегося рефлекса и более сложной реакции. Лимбическая система обусловливает эмоциональный настрой и побуждение к действию, а также процессы научения и памяти. Лимбика придает информации от внутренней среды и окружающего мира то особое значение, которое она имеет для каждого человека и тем самым определяет его целенаправленную деятельность.
Электрические раздражения различных участков лимбической системы через вживленные электроды (в эксперименте на животных и в клинике в процессе лечения больных) выявили наличие центров удовольствия, формирующих положительные эмоции, и центров неудовольствия, формирующих отрицательные эмоции. Изолированное раздражение таких точек в глубоких структурах мозга человека вызывало появление чувства «беспричинной радости», «беспредметной тоски», «безотчетного страха».
Кора больших полушарий:
Общий план организации коры. Кора больших полушарий является высшим отделом центральной нервной системы, который в процессе филогенетического развития появляется позже всего и формируется в ходе индивидуального (онтогенетического) развития позже других отделов мозга. Кора представляет собой слой серого вещества толщиной 2-3 мм, содержащий в среднем около 14 млрд. (от 10 до 18 млрд.) нервных клеток, нервные волокна и межуточную ткань (нейроглию). На поперечном ее срезе по расположению нейронов и их связей различают 6 горизонтальных слоев. Благодаря многочисленным извилинам и бороздам площадь поверхности коры достигает 0,2 м 2 . Непосредственно под корой находится белое вещество, состоящее из нервных волокон, которые передают возбуждение в кору и из нее, а также от одних участков коры другим.
Корковые нейроны и их связи. Несмотря на огромное число нейронов в коре, известно очень немного их разновидностей. Основными типами их являются пирамидные и звездчатые нейроны. В афферентной функции коры и в процессах переключения возбуждения на соседние нейроны основная роль принадлежит звездчатым нейронам. Они составляют у человека более половины всех клеток коры. Эти клетки имеют короткие ветвящиеся аксоны, не выходящие за пределы серого вещества коры, и короткие ветвящиеся дендриты. Звездчатые нейроны участвуют в процессах восприятия раздражении и объединении деятельности различных пирамидных нейронов.
Пирамидные нейроны осуществляют эфферентную функцию коры и внутрикорковые процессы взаимодействия между удаленными друг от друга нейронами. Они делятся на крупные пирамиды, от которых начинаются проекционные, или эфферентные, пути к подкорковым образованиям, и мелкие пирамиды, образующие ассоциативные пути к другим отделам коры. Наиболее крупные пирамидные клетки - гигантские пирамиды Беца - находятся в передней центральной извилине, в так называемой моторной зоне коры. Характерная особенность крупных пирамид - их вертикальная ориентация в толще коры. От тела клетки вертикально вверх к поверхности коры направлен наиболее толстый (верхушечный) дендрит, через который в клетку поступают различные афферентные влияния от других нейронов, а вертикально вниз отходит эфферентный отросток - аксон.
Многочисленность контактов (например, только на дендритах крупной пирамиды их насчитывают от 2 до 5 тыс.) обеспечивает возможность широкой регуляции деятельности пирамидных клеток со стороны множества других нейронов. Это позволяет координировать ответные реакции коры (в первую очередь ее моторную функцию) с разнообразными воздействиями из внешней среды и внутренней среды организма.
Для коры больших полушарий характерно обилие межнейронных связей. По мере развития мозга человека после его рождения увеличивается число межцентральных взаимосвязей, особенно интенсивно до 18 лет.
Функциональной единицей коры является вертикальная колонка взаимосвязанных нейронов. Вытянутые по вертикали крупные пирамидные клетки с расположенными над ними и под ними нейронами образуют функциональные объединения нейронов. Все нейроны вертикальной колонки отвечают на одно и то же афферентное раздражение (от одного и того же рецептора) одинаковой реакцией и совместно формируют эфферентные ответы пирамидных нейронов.
Распространение возбуждения в поперечном направлении-от одной вертикальной колонки к другой - ограничено процессами торможения. Возникновение активности в вертикальной колонке приводит к возбуждению спинальных мотонейронов и сокращению связанных с ними мышц. Этот путь используется, в частности, при произвольном управлении движениями конечностей.
Первичные, вторичные и третичные поля коры. Особенности строения и функционального значения отдельных участков коры позволяют выделить отдельные корковые поля.
Различают три основные группы полей в коре: сенсорные, ассоциативные и моторные поля.
Сенсорные поля связаны с органами чувств и органами движения на периферии, они раньше других созревают в онтогенезе, имеют наиболее крупные клетки. Это так называемые ядерные зоны анализаторов, по И. П. Павлову (например, поле болевой, температурной, тактильной и мышечно-суставной чувствительности находится в задней центральной извилине коры, зрительное поле(пол 17 и 18) в затылочной области, слуховое поле(поле 41) в височной области и двигательное поле(поле 6) в передней центральной извилине коры. Эти поля осуществляют анализ отдельных раздражений, поступающих в кору от соответствующих рецепторов. При разрушении сенсорных полей возникают так называемая корковая слепота, корковая глухота и т. п. Рядом расположены ассоциативные поля, которые связаны с отдельными органами только через сенсорные зоны. Они служат для обобщения и дальнейшей обработки поступающей информации. Отдельные ощущения синтезируются в них в комплексы, обусловливающие процессы восприятия. При поражении ассоциативных зон сохраняется способность видеть предметы, слышать звуки, но человек их не узнает, не помнит их значения. Сенсорные и ассоциативные поля имеются и у человека, и у животных.
Наиболее далеки от непосредственных связей с периферией третичные поля, или зоны перекрытия анализаторов. Эти поля есть только у человека. Они занимают почти половину территории коры и имеют обширные связи с другими отделами коры и с неспецифическими системами мозга. В этих полях преобладают наиболее мелкие и разнообразные клетки. Основным клеточным элементом здесь являются звездчатые нейроны. Третичные поля находятся в задней половине коры - на границах теменных, височных и затылочных ее областей и в передней половине - в передних частях лобных областей. В этих зонах оканчивается наибольшее число нервных волокон, соединяющих левое и правое полушария, поэтому роль их особенно велика в организации согласованной работы обоих полушарий. Третичные поля созревают у человека позже других корковых полей, они осуществляют наиболее сложные функции коры. Здесь происходят процессы высшего анализа и синтеза. В третичных полях на основе синтеза всех афферентных раздражении и с Учетом следов прежних раздражении вырабатываются цели и задачи поведения. Согласно им происходит программирование двигательной деятельности. Развитие третичных полей у человека связывают с функцией речи. Мышление (внутренняя речь) возможно только при совместной деятельности анализаторов, объединение информации от которых происходит в третичных полях. Разделение нейронов коры на поля, области и зоны называется функциональной мозаикой. Автором такого разделения является Бродман.
При врожденном недоразвитии третичных полей человек не в состоянии овладеть речью (произносит лишь бессмысленные звуки) и даже простейшими двигательными навыками (не может одеваться, пользоваться орудиями труда и т. п.).
Воспринимая и оценивая все сигналы из внутренней и внешней среды, кора больших полушарий осуществляет высшую регуляцию всех двигательных и эмоционально-вегетативных реакций.
Функции коры больших полушарий.
Кора больших полушарий выполняет наиболее сложные функции организации приспособительного поведения организма во внешней среде. Это прежде всего функция высшего анализа и синтеза всех афферентных раздражении.
Афферентные сигналы поступают в кору по разным каналам, в разные ядерные зоны анализаторов (первичные поля), а затем синтезируются во вторичных и третичных полях, благодаря деятельности которых создается целостное восприятие внешнего мира. Этот синтез лежит в основе сложных психических процессов восприятия, представления, мышления. Кора больших полушарий представляет собою орган, тесно связанный с возникновением у человека сознания и регуляцией его общественного поведения. Важной стороной деятельности коры больших полушарий является замыкательная функция - образование новых рефлексов и их систем (условные рефлексы, динамические стереотипы).
Благодаря необычайно большой продолжительности сохранения в коре следов прежних раздражений (памяти) в ней накапливается огромный объем информации. Это имеет большое значение для сохранения индивидуального опыта, который используется по мере необходимости.
Несмотря на анатомическую одинаковость обоих полушарий переднего мозга они функционально отличаются. Восходящие и нисходящие пути от головного мозга переходят на противоположную половину тела и поэтому левое полушарие отвечает за соматическую чувствительность и движения правой половины тела и наоборот. Также вследствие перекреста зрительных путей правая половина зрительного поля проецируется в левое полушарие, а левая половина – в правое. Изолированное правое полушарие обладает памятью, способностью к зрительному или тактильному распознаванию предметов, абстрактному мышлению и к слабому пониманию речи(выполнение слуховых команд и чтение простейших слов). В правом полушарии лучше развиты: распознавание лиц, пространственное построение и восприятие музыки. Левое полушарие является доминантным по отношению к правому. Оно обеспечивает речь и сознание, вербально – рассудочную деятельность, временные характеристики и связи событий. При его повреждении страдает логическое смысловое мышление.
Электрическая активность коры больших полушарии. Изменения функционального состояния коры отражаются на характере ее биопотенциалов. Регистрация электроэнцефалограммы (ЭЭГ), т. е. электрической активности коры, производится непосредственно с ее обнаженной поверхности (в опытах на животных и при операциях на человеке) или через неповрежденные покровы головы (в естественных условиях на животных и человеке Современные электроэнцефалографы усиливают эти потенциалы в 2-3 млн. раз и дают возможность исследовать ЭЭГ от многих точек коры одновременно.
В ЭЭГ различают определенные диапазоны частот, называемые ритмами ЭЭГ. В состоянии относительного покоя чаще всего регистрируется альфа-ритм (8-12 колебаний в 1 сек.), в состоянии активного внимания - бета-ритм (выше 13 колебаний в 1 сек.), при засыпании, некоторых эмоциональных состояниях - тэта-ритм (4-7 колебаний в 1 сек.), при глубоком сне, потере сознания, наркозе - дельта-ритм (1-3 колебания в 1 сек.).
В ЭЭГ отражаются особенности взаимодействия корковых нейронов при умственной и физической работе. Отсутствие налаженной координации при выполнении непривычной или тяжелой работы приводит к так называемой десинхронизации ЭЭГ - быстрой асинхронной активности. По мере формирования двигательного навыка происходит сонастраивание активности отдельных связанных с данным движением нейронов и отключение посторонних.
Несмотря на совершенство процессов координации в спинном мозгу, он находится под постоянным контролем головного мозга, в первую очередь коры больших полушарий.
В организме имеются специальные механизмы, обусловливающие преимущественное воздействие коры больших полушарий на общие конечные пути к мышцам-спинальные мотонейроны. Большая эффективность кортико-спинальных влияний по сравнению с сегментарными афферентными влияниями обеспечивается, во-первых, наличием прямых путей из коры к мотонейронам спинного мозга и, во-вторых, возможностью особенно быстрой их активации корковыми импульсами. Электрофизиологическими исследованиями показано, что ритмические воздействия из двигательной области коры вызывают чрезвычайно резкое нарастание суммарной амплитуды возбуждающих постсинаптических потенциалов спинальных мотонейронов. Амплитуда каждого последующего возбуждающего постсинаптического потенциала увеличивается примерно в 6 раз больше, чем при поступлении к тем же мотонейронам импульсов от проприорецепторов по афферентным путям. Таким образом, достаточно уже 2-3 импульсов, идущих от коры, чтобы деполяризация в мотонейроне достигла порогового уровня, необходимого для возникновения ответного разряда в скелетную мышцу. В результате кора больших полушарий может вызывать двигательные действия быстрее, чем периферические раздражения, и часто даже вопреки им.
В коре больших полушарий происходит выработка цели и задачи движений, соответственно этому строится и программа конкретных действий, которые нужны человеку для осуществления цели. В сложные поведенческие акты включаются не только моторные компоненты, но и необходимые вегетативные компоненты. Еще до начала движении кора больших полушарий повышает активность тех вставочных и моторных нейронов спинного мозга, которым предстоит участвовать в движении. В предстартовый период перед началом циклических движений в электрической активности коры происходит настройка на темп предстоящих движений. В тот момент, когда производится движение, кора тормозит деятельность всех посторонних афферентных путей и оказывается особенно восприимчивой к сигналам от рецепторов мышц, сухожилий и суставных сумок.
В организации двигательного акта участвуют самые различные отделы коры больших полушарий. Моторная зона коры (поле 4) посылает импульсы к отдельным мышцам, преимущественно к дистальным мышцам конечностей. Объединение отдельных элементов движения в целостный акт осуществляют вторичные поля (6-е и 8-е) премоторной области. Они определяют последовательность двигательных актов, формируют ритмические серии движении, регулируют тонус мышц. Задняя центральная извилина коры - общечувствительная область-обеспечивает субъективное ощущение движения. Здесь имеются нейроны, сигнализирующие только о возникновении движений в суставе, и нейроны, постоянно информирующие мозг о положении конечности (нейроны движения и нейроны положения).
К пространственной организации движений прямое отношение имеют задние третичные поля - нижнетеменные и теменно-затылочно-височные области коры. С их участием производится оценка удаленности и расположения предметов, оценка расположения отдельных частей собственного тела в пространстве и др. При поражении этих областей у человека теряется представление о «схеме тела» (о том, где находится нос, глаз, ухо, предплечье, спина, как опустить, например, «руки по швам»). Нарушается также представление о «схеме пространства», пространственная ориентация движения. Трудности возникают при выполнении самых простых актов: человек видит стул и узнает его, но садится мимо него; он не понимает, откуда идет звук, что означает «влево», «вправо» «вперед», «назад», не может правильно есть (например, ложка с супом попадает мимо рта) и т. д. Становится невозможным использование каких-либо орудий для трудовой или спортивной деятельности.
В высшей регуляции произвольных движений важнейшая роль принадлежит лобным долям. В третичных полях лобной коры про. исходит сознательное программирование произвольных движений определение цели поведения, двигательных задач и необходимых для их выполнения двигательных актов, а также сопоставление намеченной программы с результатами ее реализации. При регуляции лобными долями движений используется вторая сигнальная система. Движения программируются в ответ на поступающие извне словесные сигналы (словесные указания тренера, спортивные команды я пр.), а также благодаря участию внешней и внутренней речи (мышления) самого человека.
©2015-2019 сайт
Все права принадлежать их авторам. Данный сайт не претендует на авторства, а предоставляет бесплатное использование.
Дата создания страницы: 2017-06-30
Кора больших полушарий головного мозга представляет собой наиболее молодое образование ЦНС. В филогенезе объем новой коры (плаща) увеличивается. Так, новая кора по отношению ко всей коре у ежа составляет 32,4%, у кролика - 56, у собаки - 84,2, а у человека - 95,9%.
Кора больших полушарий состоит из трех зон: древней, старой и новой. В древнюю кору входят обонятельная доля, боковая обонятельная извилина. Старая кора состоит из гиппокамповой и зубчатой извилин. Новая кора представляет собой зону проекции внешней рецепции на поле воспринимаемых нейронов коры. У человека поверхность новой коры составляет 1500 см 3 . Быстрое развитие проекционных полей, ассоциативных областей коры, и медленное развитие костей черепа привело к образованию складок: борозд и извилин.
Кора состоит из 14 млрд клеток, расположенных в шести слоях (рис. 3.11).
- 1. Молекулярный слой коры головного мозга - образован волокнами, сплетенными между собой, содержит мало клеток.
- 2. Наружный зернистый слой коры головного мозга - характеризуется густым расположением мелких нейронов самой различной формы.
- 3. Наружный пирамидный слой коры головного мозга - состоит в основном из пирамидных нейронов разной величины, более крупные клетки лежат более глубоко.
- 4. Внутренний зернистый слой коры головного мозга - характеризуется рыхлым расположением мелких нейронов различной величины, мимо которых проходят плотные пучки волокон перпендикулярно к поверхности коры.
- 5. Внутренний пирамидный слой коры головного мозга - состоит в основном из средних и больших пирамидных нейронов, апикальные дендриты которых простираются до молекулярного слоя.
- 6. Слой веретеновидных клеток коры головного мозга - в нем расположены веретеновидные нейроны, глубинная часть этого слоя переходит в белое вещество головного мозга. Слои 2, 4 и 6 состоят из воспринимающих клеток. Слои 3 и 5 - пирамидные, дающие начало нисходящим двигательным путям. Восходящий путь проходит через все корковые слои (специфический путь). Неспецифический путь также проходит через все корковые слои.
Как показал киевский анатом В.А. Бец, не только вид нервных клеток, но и их взаиморасположение неодинаково в различных участках коры. Распределение нервных клеток в коре обозначается термином «цитоархитектоника». Исследования, проведенные учеными разных стран в конце XIX и начале XX столетия, позволили
Рис. 3.11.
С.Г. Кривощеков, 2012)
создать цитоархитектонические карты коры большого мозга человека и животных, в основу которых были положены особенности строения коры в каждом участке полушария. К. Бродман выделил в коре 52 цитоархитектонических поля, Ф. Фогт и О. Фогт с учетом волоконного строения описали в коре большого мозга 150 миелоар- хитектонических участков.
Белое вещество полушарий расположено между корой и базальными ганглиями. Оно состоит из большого количества волокон, идущих в разных направлениях. Это проводящие пути конечного мозга. Различают три вида этих путей:
- 1) проекционный путь. Он связывает кору с промежуточным мозгом и другими отделами ЦНС. Это восходящие и нисходящие пути;
- 2) комиссуральный путь. Его волокна входят в состав мозговых комиссур, которые соединяют соответствующие части правого и левого полушарий. Входят в состав мозолистого тела;
- 3) ассоциативные пути связывают участки коры одного и того же полушария.
В коре больших полушарий располагаются высшие регуляторные центры, обеспечивающие контроль и регуляцию всех рефлекторных процессов организма, психическую деятельность, поведение, восприятие всех видов чувствительности.
Электрическая активность коры больших полушарий. Изменения функционального состояния коры отражаются на характере ее биопотенциалов. Регистрацию электроэнцефалограммы (ЭЭГ), т.е. электрической активности коры, можно производить через неповрежденные покровы головы (в естественных условиях на животных и человеке) и регистрировать суммарную активность всех ближайших к поверхности нейронов. Современные элекгроэнцефалографы усиливают эти потенциалы в 2-3 млн раз и дают возможность исследовать ЭЭГ от многих точек коры одновременно.
В ЭЭГ различают определенные диапазоны частот, называемые ритмами ЭЭГ, и амплитуды волн (рис. 3.12). В состоянии относительного покоя чаще всего регистрируется альфа-ритм, в состоянии активного внимания - бета-ритм, при засыпании, некоторых эмоциональных состояниях - тэта-ритм, при глубоком сне, потере сознания, наркозе - дельта-ритм.
В ЭЭГ отражаются особенности взаимодействия корковых нейронов при умственной и физической работе. Отсутствие налаженной координации при выполнении непривычной или тяжелой работы приводит к так называемой десинхронизации ЭЭГ - быстрой асинхронной активности.
В процессе обучения и овладения разными двигательными навыками происходит перестройка и совершенствование функций коры больших полушарий: увеличиваются амплитуда и регулярность проявления фоновой активности - альфа-ритма в состоянии покоя, значительно усиливается по сравнению с состоянием относительного покоя взаимосвязанность (синхронность и синфазность) электриче-

Рис. 3.12. Биопотенциалы коры больших полушарий на ЭЭГ (Дж. Хэссет, 1981) ской активности различных областей коры. Это облегчает функциональные взаимодействия между различными корковыми центрами. Между этими зонами устанавливается общий ритм активности. В такие характерные системы взаимодействующих корковых зон включаются не только первичные поля (моторные, зрительные и др.), но и вторичные (например, премоторные и др.) и особенно третичные поля: передние - программирующие лобные области и задние - зоны афферентного синтеза (нижнетеменные и др.).